And... finally provides a reason for the consolidation of memory in human beings.
You'll recall that short term memory is eventually consolidated into long term memory.
And, you'll remember the earring model, where the outer hoops represent long time constants and the inner hoops go all the way to dt.
And you'll remember all the noise I made about predictive coding.
Well, look here - look what happens while memories are being consolidated:
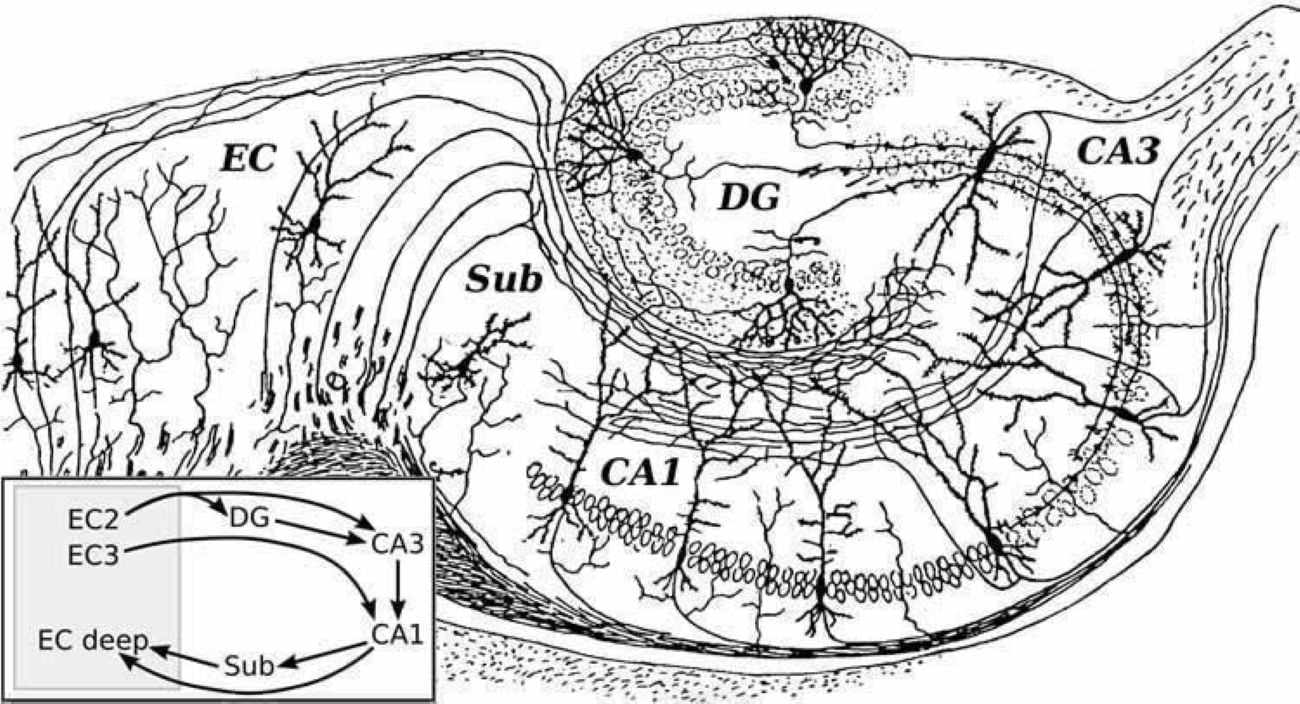
 neurosciencenews.com
neurosciencenews.com
According to the earring model, the memories are moving straight up, along a line segment connecting all the points at infinity.

And you'll recall I said two very specific things, predicted by this model:
1. Memory lives at the point at infinity. Because this is where the ends of the compactified timeline meet. Memory is the ultimate destination of all information, and the ultimate source of all information. The only way a sensory configuration turns into a motor action is by going "through" the point at infinity.
2. Awareness lives slightly ahead of "now". It's a prediction, is what it is. A prediction in the limit as dt => 0. It is a prediction that is mapped into the timeline, so it can be treated just like any other piece of information.
The amazing speed (resolution) of the network is what makes it work. Calculated at .05 picoseconds for 20.billion neurons, 5 ps if we only do cortical mini-columns, and a fraction of a nanosecond if we take full cortical columns.
That thing that you experience to be about a second wide, isn't. It's a critical state that lasts a few nanoseconds at most. The smoothness around state changes is due to the asynchronous nature of the updates, and is supported by an embedding into a sea of relatively long time constants.
The areas that glue the compactified timeline together are exactly the hippocampus and the dorsolateral prefrontal cortex - the two areas most closely related to memory.
And guess what? This also explains why phase coding is necessary! It's the only way to represent an entire pattern of activity along the timeline, as a single spike train in a single neuron.
And it also explains the sparse coding in the hippocampus.
I think, ladies and gentlemen, we have a winner. Now that the underlying mechanism is established, we can turn to more fundamental questions like "why is red red" and where am "I".
I'm pretty sure this last piece of evidence clinches the deal on the model. I'm not aware of any contradictory evidence. (Like, "any").
It makes perfect sense, too. The outer hoop is the width of the whole network, maybe a second in each direction from "now". It corresponds with "biological time frames", in other words this is how fast signals typically change when you're, say, pursuing prey or trying not to get eaten. To access that as quickly as possible the information has to move down into the inner hoops where the radius is narrower, and there, at the end of the journey in the limit as dt => 0, you get molecular memory which equates with electrical memory.
You'll recall that short term memory is eventually consolidated into long term memory.
And, you'll remember the earring model, where the outer hoops represent long time constants and the inner hoops go all the way to dt.
And you'll remember all the noise I made about predictive coding.
Well, look here - look what happens while memories are being consolidated:
Hippocampus Predicts Rewards by Reorganizing Memories - Neuroscience News
A new study finds the hippocampus reorganizes memories to predict rewards. This discovery explains how the brain learns and why Alzheimer's affects decision-making.
According to the earring model, the memories are moving straight up, along a line segment connecting all the points at infinity.
And you'll recall I said two very specific things, predicted by this model:
1. Memory lives at the point at infinity. Because this is where the ends of the compactified timeline meet. Memory is the ultimate destination of all information, and the ultimate source of all information. The only way a sensory configuration turns into a motor action is by going "through" the point at infinity.
2. Awareness lives slightly ahead of "now". It's a prediction, is what it is. A prediction in the limit as dt => 0. It is a prediction that is mapped into the timeline, so it can be treated just like any other piece of information.
The amazing speed (resolution) of the network is what makes it work. Calculated at .05 picoseconds for 20.billion neurons, 5 ps if we only do cortical mini-columns, and a fraction of a nanosecond if we take full cortical columns.
That thing that you experience to be about a second wide, isn't. It's a critical state that lasts a few nanoseconds at most. The smoothness around state changes is due to the asynchronous nature of the updates, and is supported by an embedding into a sea of relatively long time constants.
The areas that glue the compactified timeline together are exactly the hippocampus and the dorsolateral prefrontal cortex - the two areas most closely related to memory.
And guess what? This also explains why phase coding is necessary! It's the only way to represent an entire pattern of activity along the timeline, as a single spike train in a single neuron.
And it also explains the sparse coding in the hippocampus.
I think, ladies and gentlemen, we have a winner. Now that the underlying mechanism is established, we can turn to more fundamental questions like "why is red red" and where am "I".
I'm pretty sure this last piece of evidence clinches the deal on the model. I'm not aware of any contradictory evidence. (Like, "any").
It makes perfect sense, too. The outer hoop is the width of the whole network, maybe a second in each direction from "now". It corresponds with "biological time frames", in other words this is how fast signals typically change when you're, say, pursuing prey or trying not to get eaten. To access that as quickly as possible the information has to move down into the inner hoops where the radius is narrower, and there, at the end of the journey in the limit as dt => 0, you get molecular memory which equates with electrical memory.



